
![]()
New York Review Books highly
regarded Collections and new Classic Series


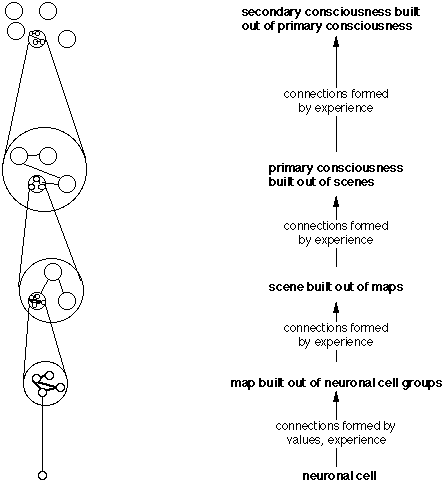
After birth a set of inborn values (feelings) (definition, more on values) allows us to begin building the structure of the brain. The smallest entity of this structure is a group of neurons (map) (definition, more on maps ) in which internal links represent our experience. Maps are then used as new building blocks and interconnected with links into scenes (definition, more on scenes) representing what we experience as the present. Ever richer maps are constructed (more on that), ultimately maps of meaning. In our search for meaning our mind develops up the evolutionary ladder to consciousness until we form the new categories of "past" and "future".
On this way, the building blocks acquire step by step more internal structure that can be accessed. A continuous stream of establishing and testing of hypotheses on the basis of the existing interconnections weakens or strengthens existing connections or builds new ones (Experiential Selection). The fittest maps and connections survive (thus the name neural Darwinism). These maps are dynamic in that they are continually redrawn (more on its definition) according to our perceptions (for more on that read the paragraphs here).
For example, disappointments or major new insights at young age may ask for major changes of the map structure and might distroy a person's drive for survival if these changes appear too radical. Similarly, works of art or psychoanalysis might strengthen some and weaken other connections in and between our maps and therewith start a re-interpretation of our perception of reality (Freud's Nachträglichkeit is an example)
At some point, the acquisition of a new kind of memory leads to a conceptual explosion. As a result, concepts of the self, the past, and the future (higher-order consciousness) can be connected to primary consciousness: selfconsciousness, culture and 'consciousness of consciousness' becomes possible.
Bright Air, Brilliant Fire is a book of astonishing variety and range, which runs from philosophy to biology to psychology to neural modeling, and attempts to synthesize them into a unified whole. It helps us understand, guide and direct our own mind (including our psyche) in that it presents a structure with which to evaluate
... The picture of psychology was a mixed one, behaviorism, gestalt psychology, psychophysics, and memory studies in normal psychology; studies of the neuroses by Freudian analysis; clinical studies of the brain lesions and motor and sensory defects ... and a growing knowledge both of neuroanatomy and the electrical behavior of nerve cells in physiology ... Only occasionally were serious efforts made ... to connect these desparate areas in a general way.Gerald Edelman, Guilio Tononi, "Consciousness: How Matter Becomes Imagination", Part III: Mechanisms of Consciousness: The Darwinian Perspective, Chapter 7: Selectionism, Degeneracy (pp. 86, 87), Penguin Books, 2000.
"All selectional systems share a remarkable property that is as unique as it is essential to their functioning: In such systems, there are typically many different ways, not necessarily structurally identical, by which a particular output occurs. We call this property degeneracy. Degeneracy is seen in quantum mechanics in certain solutions of the Schrödinger equation and in the genetic code, where, because of the degenerate third position in triplet code words, many different DNA sequences can specify the same protein.
Put briefly, degeneracy is reflected in the capacity of structurally different components to yield similar outputs or results. In a selectional nervous system, with its enormous repertoire of variant neural circuits even within one brain area, degeneracy is inevitable. Without it, a selectional system, no matter how rich its diversity, would rapidly fail -
There are countless examples of degeneracy in the brain. The complex meshwork of connections in the thalamocortical system assures that a large number of different neuronal groups can similarly affect, in one way or another, the output of a given subset of neurons. For example, a large number of different brain circuits can lead to the same motor output or action. Localized brain lesions often reveal alternative pathways that are capable of generating similar behaviors. Therefore, a manifest consequence of degeneracy in the nervous system is that certain neurological lesions may often appear to have little effect, at least within a familiar envirnoment. Degeneracy also appears at the cellular level. Neural signaling mechanisms utilize a great variety of transmitters, receptors, enzymes, and so-called second messengers. The same changes in gene expression can be brought about by different combinations of these biochemical elements.
Degeneracy is not just a useful feature of selectional systems; it is also an unavoidable consequence of selectional mechanisms. Evolutionary selective pressure is typically applied to individuals at the end of a long series of complex events. These events involve many interacting elements or muliple temporal and spatial scales, and it is unlikely that well-defined functions can be neatly assigned to independent subsets of elements or processes in biological networks. For example, if selection occurs for our ability to walk in a particular way, connections within and among many different brain structures and to the muscoleskeletal apparatus are all likely to be modified over time. While locomotion will be affected, many other functions, including our ability to stand or jump, will also be influenced as a result of the degeneracy of neural circuits. The ability of natural selection to give rise to a large number of nonidentical structures yielding simialr functions increases both the robustness of biological networks and their adaptability to unforseen environments."
Degeneracy
Degeneracy can operate at one level of organization or across many. It is seen
Edelman then began to study the nervous system, to see
The answer, Edelman proposes, is that an evolutionary process takes place - not one that selects organisms and takes millions of years, but one that occurs within each particular organism and during its lifetime, by competition among cell groups in the brain. This for Edelman is "somatic selection"....
Clearly there are some innate biases or dispositions at work; otherwise the infant would have no tendencies whatever, would not be moved to do anything, seek anything, to stay alive. These basic biases are among the first "values" we have, as Edelman calls them.
Values -simple drives, instincts, intentionalities- serve as the tools we need for adaptation and survival,
at an elementary physiological level, there are various sensory and motor "givens",Such a neuro-evolutionary view is highly consistent with some of the conclusions of psychoanalysis and developmental psychology - in particular the psychoanalyst David Stern's description of "an emergent self".We have such a certain amount of basic equipment; but, in Edelman's view, very little else is programmed or built in. It is up to the infant animal, given its elementary physiological capacities, and given its inborn values,
- from the reflexes that automatically occur (for example, in responses to pain)
- to innate mechanisms in the brain, as, for example, the feature detectors in the visual cortex which, as soon as they are activated, detect verticals, horizontals, boundaries, angles, etc., in the visual world.
- to create its own categories and
- to use them to make sense of, to construct , a world - and it is not just a world that the infant constructs, but its own world, a world constituted from the first by personal meaning and reference.
"Infants seek sensory stimulation" writes Stern. "They have distinct biases or preferences with regard to the sensations they seek. ... These are innate. From birth on, there appears to be a central tendency to form and test hypotheses about what is occurring in the world .. (to) categorize ... into conforming and contrasting patterns, events, sets, and experiences."Stern emphasizes how crucial are the active processes of connecting, correlating, and categorizing information, and how with these a distinctive organization emerges, which is experienced by the infant as the sense of a self.
Thus experience itself is not passive, a matter of "impressions" or "sensedata", but active, and constructed by the organism from the start. Active experience "selects", or carves out, a new, more complexely connected pattern of neuronal groups, a neuronal reflection of the child's individual experience, of the procedures by which it has come to categorize reality.
But these neuronal circuits are still at a low level - how do they connect with the inner life, the mind, the behavior of the creature? It is at this point that Edelman introduces the most radical of his concepts - the concepts of
But where perception of objects is concerned, the world, Edelman likes to say, is not "labeled", it does not come "already parsed (divided) into objects". We must make the objects, in effect, through our own categorizations: "Perception makes", Emerson said. "Every perception", says Edelman, echoing Emerson, "is an act of creation." In other words: our sense organs, as we move about, take samplings of the world, creating maps in the brain. Then a sort of neuronal "survival of the fittest" occurs, a selective strengthening of those mappings which correspond to "successful" perceptions - successful in that they prove the most useful and powerful for the building of "reality".
In this view, there are no innate mechanisms for complex "personal" recognition, such as the "grandmother cell" postulated by researchers in the 1970's to correspond to one's perception of one's grandmother. Nor is there any "master area", or "final common path", whereby all perceptions relating (say) to one's grandmother converge in one single place. There is no such a place in the brain where a final image is synthesized, nor any miniature person or homunculus to view this image. Rather, the perception of a grandmother or, say, of a chair depends on the synchronization of a number of scattered mappings throughout the visual cortex - mappings relating to many different perceptual aspects of the chair (its size, its shape, its color, its "leggedness", its relation to other sorts of chairs - armchairs, keeling chairs, baby chairs, etc.). In this way the brain, the creature, achieves a rich and flexible percept of "chairhood", which allows the recognition of innumerable sorts of chairs as chairs (computers, by contrast, with their need for unambiguous definitions and criteria, are quite unable to achieve this).
This perceptual generalization is dynamic, i.e. can change with time. Information coded into maps
These extensive connections allow what Edelman calls "re-entrant signaling", meaning
It must be emphasized once again:
This construct of an object or a part of reality,This re-entrant signaling is more than a feedback process that corrects errors. (Such simple feedback loops are common both in the technological world, as thermostats, governors, cruise controls, etc., and in the nervous system, where they control all of the body's automatic functions, such as temperature, blood pressure and fine control of movement.) At higher levels, where flexibility and individuality are all-important, and where new powers and new functions are needed and created, one requires "re-entrant signaling" to be a mechanism capable of constructing, not just controlling or correcting.The outputs of innumerable maps, connected by re-entry, not only complement one another at a perceptual level but are built up to higher and higher levels. For the brain, in Edelman's vision, makes maps of its own maps, or "categorizes its own categorizations", and does so by a process which can ascend indefinitely to yield ever more generalized pictures of the world.
- is not comparable to a single static image or representation -
- it is, rather, comparable to a giant and continually modulating equation.
Edelman, who himself once planned to be a concert violinist, uses musical metaphors here. "Think", he said in a recent BBC radio broadcast, "if you had a hundred thousand wires connecting four string quartet players and that, even though they weren't speaking words, signals were going back and forth in all kinds of hidden ways (as you usually get them by the subtle nonverbal interactions between the players) that make the whole set of sounds a unified ensemble. That's how the maps in the brain work by re-entry."
The players are connected. Each player, interpreting the music individually, constantly modulates and is modulated by the others. There is no final or "master" interpretation - the music is collectively created. (Gerald Edelman on re-entrant signaling, lecture given in 2006)
This, then, is Edelman's picture of the brain,
an orchestra, an ensemble - but without a conductor, an orchestra which makes its own music.Thus, two basic operations
An example: If pigeons are presented with photographs of trees, or oak leaves, or fish, surrounded by extraneous features, they rapidly learn to "home in" upon these, and to generalize, so that they can thereafter recognize any trees, or oak leaves, or fish straightaway, however distracting or confusing the context may be.
It is clear from these experiments that perception selects, or rather creates, "defining" features (what counts as "defining" may be different for each pigeon), and cognitive categories, without the use of language, or being "told" what to do. Such category-creating behavior (which Edelman calls "noetic") is very different from the rigid, algorithmic procedures used by robots. (These experiments with pigeons are described in detail in Neural Darwinism, pp. 247-251.)
Perceptual categorization, whether of colors, movements, or shapes, is the first step, and it is crucial for learning, but it is not something fixed, something that occurs once and for all. On the contrary - and this is central to the dynamic picture presented by Edelman - there is then a continual re-categorization, and this itself constitutes memory.
"In computers", Edelman writes, "memory depends on the specification and storage of bits of coded information." This is not the case in the nervous system. Memory in living organisms by contrast takes place through activity and continual re-categorization. "By its nature, memory . . . involves continual motor activity.... in different contexts.
Edelman proposed that the ability to create scenes in the mind depends upon the emergence of a new neuronal circuit during evolution, a circuit allowing for continual re-entrant signaling between,
The "scene", one must stress, is
Primary consciousness (Edelman concludes) is required for the evolution of higher-order consciousness. But it is limited -like our consciousness in a dream- to a small memorial interval around a time chunk I call the present.
Higher-order consciousness
The most difficult and tantalizing portions of Bright Air, Brilliant Fire are about how this higher-order consciousness is achieved and how it emerges from the primary consciousness. No other theorist I know of has even attempted a biological understanding of this step. To become conscious of being conscious, Edelman stresses, systems of memory must be related to representation of a self. This is not possible unless the contents, the "scenes," of primary consciousness are subjected to a further process and are themselves re-categorized.
Though language, in Edelman's view, is not crucial for the development of higher-order consciousness - there is some evidence of higher-order consciousness and self-consciousness in apes - it immensely facilitates and expands this by making possible previously unattainable conceptual and symbolic powers.
Thus two steps, two re-entrant processes, are envisaged here:
"The acquisition of a new kind of memory," Edelman writes, "...leads to a conceptual explosion. As a result, concepts of the self, the past, and the future can be connected to primary consciousness. 'Consciousness of consciousness' becomes possible."At this point Edelman makes explicit what is implicit throughout his work - the interaction of "neural Darwinism" with classical Darwinism. What occurs "explosively" in individual development must have been equally critical in evolutionary development.
Thus "at some transcendent moment in evolution", Edelman writes, there emerged "a variant with a re-entrant circuit linking value-category memory" to current perceptions". "At that moment", Edelman continues, "a memory became the substrate and servant of consciousness." And then, at another transcendent moment, by another, higher turn of re-enry, higher-order consciousness arose.There is indeed much paleontologial evidence that higher order consciousness developed in an astonishingly short space of time - some tens (perhaps hundreds) of thousands of years, not the many millions usually needed for evolutionary change. The speed of this development has always been a most formidable challenge for evolutionary theorists - Darwin himself could offer no detailed account of it and Wallace was driven back to thoughts of a grand design. But Edelman, drawing from his own observations of cell and tissue development detailed in his earlier book Topobiology, is able to suggest how it might have come about.
The principles underlying brain development and the mechanisms outlined in the Theory of Neuronal Group Selection can, he argues, account for this rapid emergence, since they allow for enormous changes in brain size over the relatively short evolutionary period in which Homo sapiens emerged. According to topobiology, relatively large changes in the structure of the brain can occur through changes in the genes that regulate the brain's morphologyn - changes that can come about as the result of relatively few mutations. And the premises of the Theory of Neuronal Group Selection allow for the rapid incorporation into existing brain structures of new and enlarged neuronal maps with a variety of functions.
This interweaving of concept and observation typifies the ambition and the grandeur of Edelman's thought. His two chapters on consciousness are the most original, the most exhilarating, and the most difficult in the entire book - but they achieve, or aspire to achieve, what no other theorist has even tried to do: a biologically plausible model of how consciousness could have emerged.
Since the Theory of Neuronal Group Selection was first formulated, important new evidence has emerged suggesting how widely separated groups of neurons in the visual cortex can become synchronized and respond in unison when an animal is faced with a new perceptual task - a finding directly suggestive of re-entrant signaling. (I discussed this work in an earlier article, "Neurology and the Soul.", New York Book Review, November 20, 1990) There is also much evidence of a more clinical sort, which one feels may be illuminated, and perhaps explained, by the Theory of Neuronal Group Selection.
I often encounter situations in day-to-day neurological practice which completely defeat classical neurological explanations, which cry out for explanations of a radically different kind, and which are clarified by Edelman's theory. (Some of these situations are discussed by Israel Rosenfield in his new book: The Strange, Familiar and Forgotten, wbere he speaks of "the bankruptcy of classical neurology".) Thus if a spinal anesthetic is given to a patient - as used to be done frequently to women in childbirth - there is not just a feeling of numbness below the waist. There is, rather, the sense that one terminates at the umbilicus, that one's corporeal self has no extension below this, and that what lies below is not-self, not-flesh, not-real, not-anything. The anesthetized lower half has a bewildering nonentity, completely lacks meaning and personal reference. The baffled mind is unable to categorize it, to relate it in any way to the self. One knows that sooner or later the anesthetic wear off, yet it is impossible to imagine the missing parts in a positive way. There is an absolute gap in primary consciousness which higher order consciousness can report, but cannot correct.
This indeed is a situation I know well from personal no less than clinical experience, for it is what I experienced in myself after a nerve injury to one leg, when for a period of two weeks, while the leg lay immobile and senseless, I found it "alien," not me, not real. I was astonished when this happened, and unassisted by my neurological knowledge - the situation was clearly neurological, but classical neurology has nothing to say about the relation of sensation to knowledge and to "self"; about how, normally, the body is "owned"; and how, if the flow of neural information is impaired, it may be lost to consciousness, and "disowned" - for it does not see consciousness as a process
Such body-image and body-ego can be fully understood, in Edelman's thinking, as breakdowns in local mapping, consequent upon nerve damage or disuse. It has been confirmed, further, in animal experiments that the mapping is not something fixed, but plastic and dynamic, and dependent upon a continual inflow of experience and use; and that if there is continuing interference with, say, one's perception of a limb or its use, there is not only a rapid loss of its cerebral map, but a rapid remapping of fhe rest of the body which then excludes the limb itself.
Stranger still are the situations which arise when the cerebral basis of body-image is affected, especially if the right hemisphere of the brain is badly damaged in its sensory areas. At such times patients may show an "anosognosia," an unawareness that anything is the matter, even though the left side of the body may be senseless, and perhaps paralyzed, too. Or they may show a strange levity, insisting that their own left sides belong to "someone else." Such patients may behave (as an eminent neurologist, M.M. Mesulam, has written) ". . . as if one half of the universe had abruptly ceased to exist . . . as if nothing were actually happening [there] . . . as if nothing of importance could be expected to occur there." Such patients live in a hemispace, a bisected world, but for them, subjectivly, their space and world is entire. Anosognosia is unintelligible (and was for years misinterpreted as a bizarre neurotic symptom) unless we see it (in Edelman's term) as a "disease of consciousness," a total breakdown of high-level re-entrant signaling and mapping in one hemisphere - the right hemisphere, which, Edelman suggests, may have only primary but no higher-order consciousness - and a radical reorganization of consciousness in consequence.
Less dramatic than these complete disappearances of self or parts of the self from consciousness, but still remarkable in the extreme, are situations in which, following a neurological lesion, a dissociation occurs between perception and consciousness, or memory and consciousness, cases in which there remains only "implicit" perception or knowledge or memory. Thus my amnesiac patient Jimmie ("The Lost Mariner") had no explicit memory of Kennedy's assassination, and would indeed say, "No president in this century has been assassinated, that I know of." But if asked, "Hypothetically, then, if a presidential assassination had somehow occurred without your knowledge, where might you guess it occurred: New York, Chicago, Dallas, New Orleans, or San Francisco?" he would invariably "guess" correctly, Dallas.
Similarly, patients with visual agnosias, like Dr. P. ("The Man who Mistook his Wife for a Hat"), while not concsciously able to recognize anyone,often "guess" the identity of peoples faces correctly. And patients with cortical blindness, from massive bilateral damage to the primary visual areas of the brain, while asserting that they can see nothing, may also mysteriously "guess" correctly what lies before them - so-called "blindsight." In all these cases, then, we find that perception, and perceptual categorization of the kind described by Edelman, has been preserved, but has been divorced from consciousness.
In such cases it appears to be only the final process, in which the re-entrant loops combine memory with current perceptual categorization, that breaks down. Their understanding, so elusive hitherto, seems to come closer with Edelman's "re-entrant" model of consciousness.
Dissatisfaction with the classical theories is not confined to clinical neurologists; it is also to be found among theorists of child development, among cognitive and experimental psychologists, among linguists, and among psychoanalysts. All find themselves in need of new models. This was abundantly clear in May of 1992, at an exciting conference on "Selectionism and the Brain" held at the Neurosciences Institute in New York and attended by prominent workers in all of these fields.
Particularly suggestive was the work of Esther Thelen and her colleagues at the University of Indiana in Bloomington, who have for some years been making a minute analysis of the development of motor skills - watching, reaching for objects - in infants. "For the developmental theorist," Thelen writes, "individual differences pose an enormous challenge.... Developmental theory has not met this challenge with much success." And this is, in part, because individual differences are seen as extraneous, whereas Thelen argues that it is precisely such differences, the huge variation between individuals, that allow the evolution of unique motor patterns.
Thelen found that the development of such skills, as Edelman's theory would suggest, follows no single programmed or prescribed pattern. Indeed there is great variability among infants at first with many patterns of reaching for objects; but there then occurs, over the course of several months, a competition among these patterns, discovery or selection of workable patterns, or workable motor solztions. These solutions, though roughly similar (for there are a limited number of ways in which an infant can reach), are always dlfferent and individual, adapted to the particular dynamics of each child, and they emerge by degrees, through exploration and trial. Each child, Thelen showed, explores a rich range of possible ways to reach for an object and selects its own path, without the benefit of any blueprint or program. The child is forced to be original, to create its own solutions. Such an adventurous course carries its own risks - the child may evolve a bad motor solution - but sooner or later such bad solutions tend to destabilize, break down, and make way for further exploration, and better solutions.
Similar considerations arise with regard to recovery and rehabilitation after strokes and other injuries.There are no rules, there is no prescribed path to recovery; every patient must discover or create his own motor and perceptual patterns, his own solutions to the challenges that face him; and it is the function of a sensitive therapist to help him in this. This is well understood in the practice of "functional integration," pioneered by Moshe Feldenkrais, and used increasingly both in rehabilitation after injury and in the training of dancers and athletes. "One cannot teach a person how to organize movement or how to perceive", writes Carl Ginsburg, a leading Feldenkrais teacher. "We need a system that organizes itself as it experiences. . . a system that has both stability and extraordinary plasticity to shift with changing circumstances. It is a system that is exceedingly difficult to model." Ginsberg feels that Theory of Neuronal Group Selection is closest to the model required ("The Roots of Functional Integration, Part III: The Shift in Thinking", The Feldenkrais Journal, No. 7 (Winter 1992), pp. 3447.
When Thelen tries to envisage the neural basis of such learning, she uses terms very similar to Edelman's: she sees a "population of movements being selected or "pruned" by experience. She writes of infants "remapping" the neuronal groups that are correlated with their movements, and "selectively strengthening particular neuronal groups". She has, of course, no direct evidence for this, and such evidence cannot be obtained until we have a way of ~visualizing vast numbers of neuronal groups simultaneously in a conscious subject, and following their interactions for months on end. No such visualization is possible at the present time, but it will perhaps become possible by the end of the decade. Meanwhile, the close correspondence between Thelen's observations and the kind of behavior that would be expected from Edelman's theory is striking.
If Esther Thelen is concerned with direct observation of the development of motor skills in the infant, Arnold Modell of Harvard, at the same conference, was concerned with psychoanalytical interpretations of early behavior; he too felt, like Thelen, that a crisis had developed, but that it might also be resolved by the Theory of Neuronal Group Selection - indeed, the title of his paper was "Neural Darwinism and a Conceptual Crisis in Psychoanalysis". The particular crisis he spoke of was connected with Freud's concept of Nachträglichkeit, the retranscription of memories which had become part of pathological fixations but were opened to consciousness, to new contexts and reconstructions, as a crucial part of the therapeutic process of liberating the patient from the past, and allowing him to experience and move freely once again.
This process cannot be understood in terms of the classical concept of memory in which a fixed record or trace or representation is stored in the brain - an entirely static or mechanical concept - but requires a concept of memory as active and "inventive" (see Israel Rosenfeld, The Invention of Memory: A New View of the Brain (Basic Books, 1991). That memory is essentially constructive (as Coleridge insisted, nearly two centuries ago) was shown experimentally by the great Cambridge psychologist Frederic Bartlett. "Remembering," he wrote, is not the re-excitation of innumerable fixed, lifeless and fragmentary traces. It is an imaginative reconstruction, or construction, built out of the relation of our attitude toward a whole mass of organized past reactions or experience.
It was just such an imaginative, context-dependent construction or reconstuction that Freud meant by Nachträglichkeit - but this, Modell emphasizes, could not be given any biological basis until Edelman's notion of memory as re-categorization. Beyond this, Modell as an analyst is concerned with the question of how the self is created, the enlargement of self through finding, or making, personal meanings. Such a form of inner growth, so different from "learning" in the usual sense, he feels, may also find its neural basis in the formation of ever-richer but always self-referential maps in the brain, and their incessant integration through re-entrant signaling, as Edelman has described it. Modell's ideas have been set out in full in Other Times, Other Realities (Harvard University Press, 1990), and in a forthcoming book, The Private Self(Harvard University Press, 1993).
Others too - cognitive psychologists and linguists - have become intensely interested in Edelman's ideas, in particular by the implication of the extended Theory of Neuronal Group Selection which suggests that the exploring child, the exploring organism, seeks (or imposes) meaning at all times, that its mappings are mappings of meaning, that its world and (if higher consciousness is present) its symbolic systems are constructed of "meanings." When Jerome Bruner and others launched the "cognitive revolution" in the mid 195Os, this was in part a reaction to behaviorism and other "isms" which denied the existence and structure of the mind. The cognitive revolution was designed "to replace the mind in nature", to see the seeking of meaning as central to the organism. In a recent book, Acts of Meaning, (Harvard University Press, 1990), Bruner describes how this original impetus was subverted, and replaced by notions of computation, information processing, etc., and by the computational (and Chomskian) notion that the syntax of a language could be separated from its semantics.
But, as Edelman writes, it is increasingly clear, from studying the natural acquisition of language in the child, and, equally, from the persistent failure of computers to "understand" language, its rich ambiguity and polysemy, that syntax cannot be separated from semantics. It is precisely through the medium of "meanings" that natural language and natural intelligence are built up. From Boole, with his "Laws of Thought" in the 1850s, to the pioneers of Artificial Intelligence at the present day, there has been a persistent notion that one may have an intelligence or a language based on pure logic without anything so messy as "meaning" being involved. That this is not the case, and cannot be the case, may now find a biological grounding in the Theory of Neuronal Group Selection.
Edelman and his colleagues at the Neurosciences Institute have been deeply interested in such "synthetic neural modeling", and have devised a series of "synthetic animals" or artifacts designed to test the Theory of Neuronal Group Selection. Although these "creatures" - which have been named DARWIN I, II, III, and IV - make use of supercomputers, their behavior (if one may use the word) is not programmed, not robotic, in the least, but (in Edelman's word) "noetic." They incorporate both a selectional system and a primitive set of "values"- for example, that light is better than no light - which generally guide behavior but do not determine it or make it predictable. Unpredictable variations are introduced in both the artifact and its enviromnent so that it is forced to create its own categorizations.
DARWIN IV or NOMAD, with its electronic eye and snout, has no "goal", no "agenda", but resides in a sort of pen, a world of varied simple objects (with different colors, shapes, textures, weights).
Her follows an illustration showing N0DMAD, an adaptive device constructed by Gerald M. Edelman and his colleagues at the Neurosience Institute, in its environment. NOMAD is controlled by a computer simulated "selectionist nervous system.". It has a TV camera "eye" and a snout with eletrical "taste" sensors. Synapses in its simulated brain change with experience so that NOMAD learns to approach and taste blocks. After forming an assosiation between taste and color, it avoids bad-tasting blocks (blue) but collects tasty ones (red).
True to its name, it wanders around like a curious infant, exploring these objects, reaching for them, classifying them, building with them, in a spontaneous and idiosyncratic way (the movement of the artifact is exceedingly slow, and one needs time-lapse photography to bring home its creatural quality). No two "individuals" show identical behavior - and the details of their reachings and learnings cannot be predicted, any more than Thelen can predict the development of her infants. If their value circuits are cut, the artifacts show no learning, na "motivation", no convergent behavior at all, but wander around in an aimlessway, like patients who have had their frontal lobes distroyed. Since the entire circuitry of these DARWINS is known, and can be seen functioning on the screen of a supercomputer, one can continuously monitor their inner workings, their internal mappings, their re-entrant signalings - one can see how they sample their environment, one can see how the first, vague, tentative peercepts emerge, and how, with hundreds of further samplings, they evolve and become recognizable, refined models of reality, following a process similar to that projected by Edelman's theory.
Normally one is not aware of the brain's almost automatic generation of "perceptual hypotheses" (in Richard Gregory's terms) and their refinement through a process of repeated samplings and testing. But under certain circumstances, as in recovery after acute nerve injury, one may become vividly aware of these normally unconscious (and sometimes exceedingly rapid) operations. I give a personal example of this in A Leg to Stand On.
Seeing the DARWINS, especially DARWIN IV, at work can induce a curious state of mind. Going to the zoo after my first sight of DARWIN IV, I found myself looking at birds, antelopes, lions, with a new eye: were they, so to speak, nature's DARWINS, somewhere up around DARWIN XII in complexity? And the gorillas, with higher-order consciousness but no language - where would they stand? DARWIN XIX? And we, writing about the gorillas, where would we stand? DARWIN XXVII perhaps? A particularly intriguing, sometimes frightening part of Bright Air, Brilliant Fire is its penultimate chapter, "Is It Possible to Construct a Conscious Artefact?" Edelman has no doubt of the possibility, but places it, mercifully, well on in the next century.
Such then is the sweep of Bright Air, Brilliant Fire, and its central ambition of "replacing the mind in nature". It is a book of astonishing variety and range, which runs from philosophy to biology to psychology to neural modeling, and attempts to synthesize them into a unified whole.
Neural Darwinism (or Neural Edelmanism, as Francis Crick has called it) coincides with our sense of "flow," that feeling we have when we are functioning optimally, of a swift, effortless, complex, ever changing, but integrated and orchestrated stream of consciousness; it coincides with the sense that this consciousness is ours, and that all we experience and do and say is, implicitly, a form of selfexpression, and that we are destined, whether we wish it or not, to a life of particularity and self-development; it coincides, finally, with our sense that life is a journey - unpredictable, full of risk and uncertainty, but, equally, full of novelty and adventure, and characterized (if not sabotaged by external constraints or pathology) by constant advance, an ever deeper exploration and understanding of the world.
Edelman's theory proposes a way of grounding all this in known facts about the nervous system and testable hypotheses about its operations. Any theory, even a wrong theory, is better than no theory; and this theory - the first truly global theory of mind and consciousness, the first biological theory of individuality and autonomy - should at least stimulate a storm of experiment and discussion.
Merlin Donald, at the end of his fine and far-reaching recent book Origins of the Modern Mind (Harvard University Press, 1991), speaks of this in his conclusion:
Mental materialism is back, with a vengeance. It is not only back, but back in an unapologetic, out-of-the-closet, almost exhibitionistic form. This latest incarnation might be called "exuberant materialism." Changeux (1985), Churchland (1986), Edelman (1987), Young (1988), and many others have announced a new neuroscientific apocalypse.
Optimism is basically more productive than pessisism, and exuberant materialists are certainly optimists. Neuroscience is in its adolescence, and the field is drunk ; with its own dizzying growth; how not to be optimistic?
There is no better place to read about this than in Edelman's own works, dense and difficult though they frequently are. Bright Air, Brilliant Fire is the most wide-ranging and accessible. It is strenuous and sometimes maddening, and one must struggle to understand it; but if one struggles, if one reads and reads again, the stubborn paragraphs finally yield their meaning, and a brilliant and captivating new vision of the mind emerges.
|
Oliver Sacks in September 2011 on Web of STORIES |
At the time he wrote this review, Oliver
Sacks was Professor of Neurology at the Albert Einstein College of
Medicine in New York. His books include Awakenings (in which he
described some of his work at Beth Abraham Hospital, the Bronx, New York
City. This is the hospital where he is now, in 1996), A Leg to Stand
On, The Man Who Mistook His Wife for a Hat, and, published
shortly before this review, Seeing Voices Recently he finished
The
Island of the Colorblind More on/of Oliver Sacks:
|